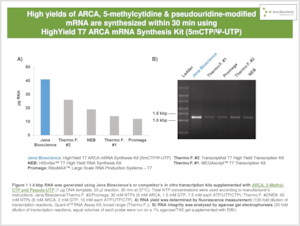
Synthetic mRNAs are widely used as alternative molecules to plasmid DNA for modulating protein levels in a therapeutic and research context. Their biological functionality depends on both a 5’ Cap structure and nucleotide modifications that ensure efficient translation and reduced immunogenicity [1]-[16]. The classic combination of modifications consists of ARCA (Cap 0), 5-methylcytidine and pseudouridine which are conveniently introduced by correspondingly modified nucleotides via in vitro transcription.
Beyond this classic combination, other mRNA modifications (e.g. N1-Methylpseudouridine [10], N4-Acetyl-Cytidine [5,6]) or the introduction of a Cap 1 moiety [15,16]) have been demonstrated to increase translation efficiency and/or to reduce immunogenicity as well. The optimal combination of Cap moiety & nucleotide modification however, needs to be individually determined for each mRNA target. Investigations can conveniently be performed with HighYield T7 mRNA Testkits.
[1] Karikó et al.(2005) Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity23:165.
[2] Karikó et al.(2008) Incorporation of Pseudouridine into mRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Mol. Ther.16(11):1833.
[3] Kormann et al.(2011) Expression of therapeutic proteins after delivery of chemically modified mRNA in mice. Nature Biotechnology29(2):154.
[4] Warren et al.(2011) Highly Efficient Reprogramming to Pluripotency and Directed Differentiation of Human Cells with Synthetic Modified mRNA. Cell Stem Cell7:618.
[5] Svitkin et al.(2017) N1-methyl-pseudouridine in mRNA enhances translation through eIF2alpha-dependent and independent mechanisms by increasing ribosome density. Nucleic Acid Res45(10):6023.
[6] Andies et al.(2015) N1-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J. Control. Release217:337.
[7] Li et al.(2016) Effects of Chemically Modified Messenger RNA on Protein Expression. Bioconjugate Chem.27:849.
[8] Arango et al.(2018) Acetylation of Cytidine in mRNA Promotes Translation Efficiency. Cell175(7):1872.
[9] Sinclair et al.(2017) Profiling Cytidine Acetylation with Specific Affinity and Reactivity. ACS Chem. Neurosci.12(12):2922.
[10] Dominissini et al.(2016) The dynamic N1-methyladenosine methylome in eukaryotic messenger RNA. Nature530:441.
[11] Wienert et al. (2018) In vitro transcribed guide RNAs trigger an innate immune response via RIG-I pathway. PLoS Biol. 16 (7) :e2005840.
[12] Kim et al. (2018) CRISPR RNAs trigger innate immune responses in human cells. Genome Res. 28 (3):367.
[13] Badieyan et al. (2019) Concise Review: Application of Chemically Modified mRNA in Cell Fate Conversion and Tissue Engineering. Stem Cells Translational Medicine8:833.
[14] Hadas et al. (2019) Optimizing Modified mRNA In Vitro Synthesis Protocol for Heart Gene Therapy. Molecular Therapy: Methods & Clinical Development 14:300.
[15] Shatkinet al. (1976) Capping of eukaryotic mRNAs. Cell 9(4):645.
[16] Gallowayet al.(2019) mRNA cap regulation in mammalian cell function and fate. Biochimica et Biophysica Acta 1862(3):270.